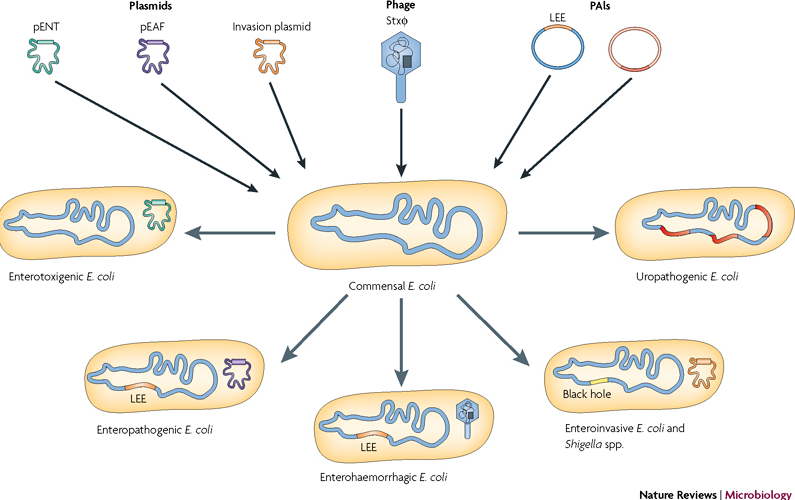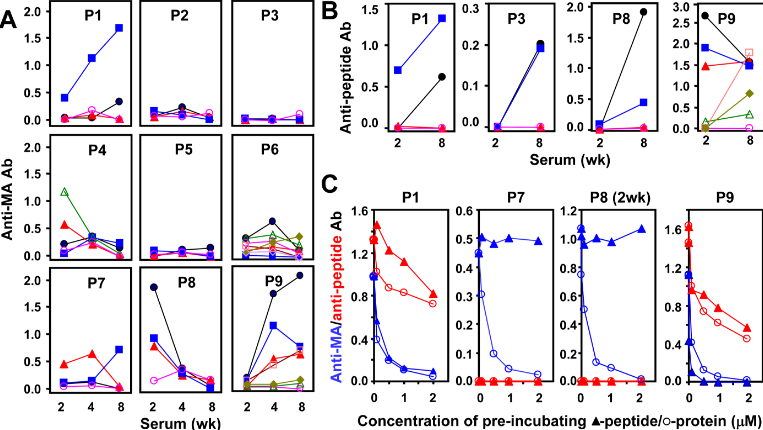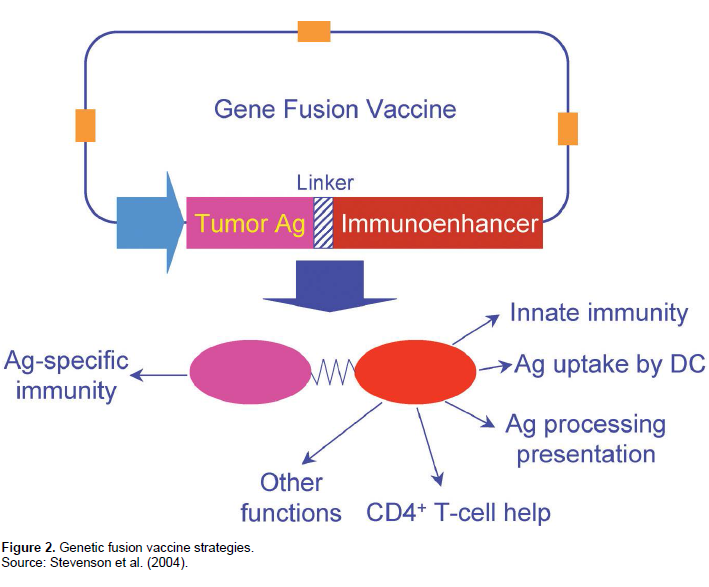
Categories : NewsLeave a comment
Well Known Schools that teaches Microbiology
Pharmaceutical and Generic Engineering firms are searching for Microbiologist. If you are in U.S and are in need of the best school that provides the best education for those who wanted to become a Microbiologist, the top 4 includes Harvard University, Stanford, University of California—Berkeley, and University of Wisconsin—Madison. These